Por: Orlando Valega , apicultor de “Apícola Don Guillermo”Correo: apicoladonguillermo@yahoo.com.ar

Cuando
seleccionamos animales o plantas por lo general lo hacemos por las
características individuales que estas presentan, ya sea por su
belleza, performance, resistencia a las enfermedades, productividad,
etc. En la abeja las características individuales dicen poco o nada. La
abeja se comporta como un organismo social formando una colonia en la
que conviven e interactúan individuos de distinto sexo y de cierto grado
variable de parentesco. Algunos la consideran como una superfamilia
integrada por varias subfamilias y otros como un superorganismo ya que
este mantiene temperatura y humedad constante además de nutrirse cada
individuo en forma mancomunada con los demás individuos de la colonia
gracias a un mecanismo denominado trofalaxia, o intercambio de
alimentos . Si la colonia pasa hambre pasan hambre todos sus integrantes
por igual.
Una colonia está integrada por una REINA MADRE
encargada de la multiplicación de los individuos. Esta mantiene in-situ
las células germinativas masculinas y femeninas apareándose en edad muy
temprana con varios machos (acoplamientos múltiples) de 10 a 17
zánganos, a los que les extirpa los órganos sexuales y retiene las
células germinativas en un depósito llamado espermateca.
Ovoposita:
a) Óvulos (haploides) de los que nacen machos normales, los ZÁNGANOS;
b) Huevos (diploides heterocigotas) de los que nacen hembras fértiles, las PRINCESAS y hembras estériles, las OBRERAS y
c) Huevos (diploides homocigotas) de los que nacen unos monstruitos medio machos y medio hembras, que son comidos antes del tercer día de nacidos por las obreras.
Las obreras integran varias subfamilias que tienen un padre y madre en común pero como el padre proviene del óvulo sin fecundar .de esta manera las obreras que pertenecen a la misma subfamilia, tienen tres cuartos de sus genes en común. Reciben gametos idénticos de su padre y, en promedio, gametos mitad-idénticos de su madre.
b) Huevos (diploides heterocigotas) de los que nacen hembras fértiles, las PRINCESAS y hembras estériles, las OBRERAS y
c) Huevos (diploides homocigotas) de los que nacen unos monstruitos medio machos y medio hembras, que son comidos antes del tercer día de nacidos por las obreras.
Las obreras integran varias subfamilias que tienen un padre y madre en común pero como el padre proviene del óvulo sin fecundar .de esta manera las obreras que pertenecen a la misma subfamilia, tienen tres cuartos de sus genes en común. Reciben gametos idénticos de su padre y, en promedio, gametos mitad-idénticos de su madre.
Las obreras que pertenecen a diversas
subfamilias tienen la misma madre pero diversos padres. Son medias
hermanas y tienen un cuarto de sus genes en común. Los Zánganos son
medio hermanos con las obreras y con la REINA
Esta gran familia o superorganismo no puede estudiarse por las características aisladas e individuales de uno de sus miembros, mas bien por las características que la misma en su conjunto refleja. Por lo tanto si los portadores de las características genéticas son las REINAS y los ZÁNGANOS juzgaríamos a la reina no por las características individuales de ella, sino por las características de la super-familia o super-organismo que esta compone y de los ZÁNGANOS sabemos que son medio hermanos de la reina.
Además de lo complicado que resulta, por las razones antedichas, la aplicación de las leyes de la genética a la apicultura, tenemos los inconvenientes derivados de la partenogénesis en las reinas; el acoplamiento múltiple; acoplamiento con zánganos del origen desconocido; el hecho de que cada zángano muere en el acto del acoplamiento y por lo tanto no se puede utilizar para otros acoplamientos como es posible en otros ámbitos productivos. Y no olvidemos recordar que la reina se aparea muy joven y por única vez, después por el resto de su vida utiliza los espermas acumulados en la espermateca.(*) Aunque la abeja está conforme a los leyes universales válidas de la genéticas según lo establecido por MENDEL, sin embargo manifiesta excepciones y particularidades de vital importancia. PARTENOGÉNESIS (MITOS SOBRE EL ZÁNGANO)
La partenogénesis (del idioma griego παρθένος parthenos = virgen + γένεσις genesis = generación) es una forma de reproducción asexual por hembras no fecundadas , que se da con cierta frecuencia en insectos, anfibios y reptiles . Fue Jan Dzierzon quien primero descubrió la partenogénesis en los zánganos. Puede también considerarse como reproducción asexual o como sexual mono-gamética, puesto que interviene en ella una célula sexual o gameto
Como se sabe, las hembras de las abejas -reinas y obreras- nacen de un huevo, que es la unión de un óvulo de la reina con un espermatozoide de uno de los 10 a 15 zánganos que se aparearon con esa reina. En tanto que los zánganos nacen de los óvulos de la reina, esto significa que la reina produce zánganos sin necesidad de aparearse. Esta particularidad del sistema reproductivo de las abejas se denominó PARTENOGENESIS en 1845 cuando Dzierzon describe por primera vez el fenómeno. En 1986 el brillante apicultor uruguayo Samuel de León del Río, lo rebautizó como TELIGENESIS, en su muy interesante libro "He aquí las abejas" (Editorial Hemisferio Sur), y nos ayudó a comprender que los zánganos de una colmena son medios hermanos de la reina que puso el óvulo de donde nacieron, y no sus hijos como la teoría de la partenogénesis establecía.
Existen muchos mitos acerca del zángano. Siempre se afirmó que el zángano era hijo de la reina, cuando la realidad es que genéticamente es hermano de la reina, pues parte de una unidad biológica a la que mal se ha hecho en denominar huevo, ya que para que fuese un huevo debería contener la información genética de dos progenitores y en realidad es un óvulo formado a partir de una meiosis de las células de la reina.
Por esa razón es incorrecto decir que el zángano nace de un huevo infecundo porque en realidad ni es un huevo ni es infecundo. Podríamos llamarla MADRE HERMANA Para poder entender esto, puede citarse el caso de la mosca de la fruta o Ceratitis capitata. Esta mosca es combatida criando machos en cautiverio a los que se esteriliza en laboratorios para que luego al ser liberados fecunden a las hembras. Como los machos no tendrán semen al fecundar las hembras, éstas pondrán huevos que jamás incubarán por falta de uno de los gametos.
Si este mismo proceso se efectuara con las abejas y se esterilizaran los zánganos, ellas igualmente pondrían "huevos" que originarían machos.
Otro de los mitos existentes es el de afirmar que los zánganos serán todos iguales a su "madre hermana" cuando en realidad lo que sucede es lo contrario, pues la célula que origina al macho, ese óvulo hasta ahora llamado huevo infecundo, surge por una meiosis celular de la reina, es decir que de los 32 cromosomas apareados que forman el genoma de la reina sólo 16 pasarán al zángano.
EFECTOS DE LA PARTENOGÉNESIS
Aparte del hecho de que tenemos en el mejor de los casos, solamente un control limitado del acoplamiento de las reinas, es la partenogénesis el verdadero punto crítico en la crianza de la abeja. La partenogénesis anula, no solamente los procesos de crianza normales, también rompe todas nuestras nociones e hipótesis preconcebidas referentes a la herencia. Debido a la partenogénesis el zángano no tiene ningún padre sino solamente una madre “Madre Hermana”. Por otra parte él pierde su vida en el acto del acoplamiento y así deja de estar de servicio para los propósitos posteriores de la crianza. Por lo tanto no hay posibilidad en la abeja de acoplamientos entre el PADRE E HIJA, MADRE E HIJO (*), o HERMANO Y HERMANA. De máxima puede haber acoplamiento emparentado entre un HERMANASTRO Y UNA HERMANASTRA.
La situación es mas complicada por el hecho que, debido a la partenogénesis, millones de espermatozoides producidos por un zángano son todos absolutamente idénticos desde el punto de vista genético.
Un resultado de esta uniformidad de genes en el zángano significa que hay una mayor estabilidad en la herencia de la abeja que en otras formas de vida. Otra consecuencia de esta uniformidad es que la abeja es más susceptible a la endogamia. Es verdad que el acoplamiento múltiple actúa como contrapeso a esto, pero solamente parcialmente. En nuestro cruce obtenemos una segregación en el descendiente femenino en la F1, en los zánganos solamente en la F2, pero aquí no hay después el mismo patrón que en otros tipos de crianzas donde no hay partenogénesis. Cruzando a individuos de la F1 entre sí mismos, MENDEL podía obtener la segregación clásica en la F2, (Fenotipo 3:1) de la cual se presentan las nuevas combinaciones de los genes que entonces se transmiten en una línea directa. Tales nuevas combinaciones son posibles en el caso de la abeja, pero como se ha observado ya, solamente de una manera del cruce giratorio debido a el hecho de la partenogénesis.
El zángano no tiene ningún hijo, solamente nietos; él no tiene padre tiene solo madre y sus factores hereditarios correspondan a los de sus abuelos. Por lo tanto en un primer cruzamiento no tenemos ningún zángano de F 1, ellos aparecen solamente en la generación subsiguiente.
En las hembras fértiles se puede producir la hibridación y la segregación de genes en el primer cruzamiento (F1) pero no así en el zángano que es un mero multiplicador y transmisor de los gametos femeninos. En el zángano se produce la segregación recién en el segundo cruce o (F2)
Los zánganos nacen de un óvulo y tienen la mitad de cromosomas 16 y sus espermas tienen también 16 cromosomas por lo tanto repite las características genéticas del óvulo que lo engendró.
Las abejas al igual que las reinas tienen dos GAMETOS y por eso pueden ser “puras de una raza” o “mestizas”. En casos excepcionales desovan!. Los zánganos sólo tienen un GAMETO, nacen de un óvulo no fecundado por eso nacen PUROS DE ALGUNA RAZA ya que para el mestizaje habría necesidad de dos GAMETOS. Puede haber algún intercambio de genes por (“CROSSING-OVER”) o Sobrecruzamiento.
La espermateca de la REINA (local donde se quedan depositados vivos los espermatozoides de los zánganos) no tiene evidentemente, por lo tanto, la mínima influencia sobre su prole de zánganos (Ha no ser en sus hijas y nietas reinas). Sabedor de eso el apicultor o Genetista podrá programar nacimientos de zánganos aunque sus REINAS se hubieran fecundado con machos de otras razas. (*). Por eso una reina ITALIANA PURA aún fecundada por ZÁNGANOS AFRICANOS generará solamente ZÁNGANOS ITALIANOS. Si la reina fuera híbrida irá a producir simultáneamente dos razas diferentes de zánganos. Como ejemplo una Reina ITALIANA AFRICANIZADA irá a dar origen a la mitad de zánganos ITALIANOS (derivados del GAMETO “A )”y la otra mitad será de AFRICANOS (oriundos del GAMETO “B”). En mestizas es muy común encontrar un grupo de zánganos bien amarillos y otros bien oscuros
Por las Leyes de MENDEL son fácilmente recuperadas las RAZAS puras de entre las cruzas (mestizos). Esto se está haciendo hace más de 70 años en los Centros de Genética y sin que hayan ocurrido mayores complicaciones. Sobreestimar el SOBRECRUZAMENTO no explicaría porque después de tantos años de mestizajes en poblaciones cerrados y en constantes depuraciones no se quedó todo emparentado a tal punto que inviabilizase los desoves de aquellas reinas.
En la F2 cruzando el Híbrido KI con un zángano K (tercero) dará hijas de de la RAZA CÁRNICA Y otra vez un mestizo pero ahora el gameto del zángano esta del lado femenino IK dando un mestizo Italiano Carnicizado
En el de la derecha (cuarto)aparecen las ITALIANAS puras (“II”) y otra vez el híbrido invertido explicando como el gameto del zángano tiene segregación en la F2. Nadie puede negar esta Ley básica de MENDEL de que de los mestizos apareados reaparecen inmediatamente las RAZAS puras que les dieron el origen.
Es evidente que en BRASIL las APIS MELLÍFICA son africanas puras porque no hay abejas, reinas y zánganos europeos en los enjambres y ni los respectivos híbridos lo que sería una inevitabilidad científica; también tampoco existe la menor posibilidad aún usando las técnicas de la Inseminación Artificial de rescatar las razas antiguas pre-existentes a las africanas A. M. SCUTELLATA. De esto sólo puede ser explicada una conclusión lógica: las europeas y sus mestizas - las africanizadas - ya no existen y lo que hoy son, sí, solamente africanas puras (“AA”) cómo lo confirmó la cuenta de genes. Claudio Mikos
Es verdad que existe un concepto adicional: el del PEDIGRI. Las hijas del primer Gráfico podrían ser Pedigris si aquella MATRIZ (“KK – K”) tuviera un CERTIFICADO DE ORIGEN. En el caso de las CÁRNICAS (“KK”) y de las ITALIANAS (“II”) del Gráfico de la derecha a pesar de que sean tenidas cómo puras evidentemente no podrían ser calificadas como Pedigris. En el comercio de material genético como lo del ganado es usado el término “PO” (“PURO DE ORIGEN”) lo cual es cotizado por elevados precios. En el caso de las abejas es poco usual probablemente debido al hecho de que actualmente es muy difícil encontrar ejemplares sin que hayan sido mezclados en el pasado con otras razas. Hay además el detalle de que las reinas se aparean lejos de las colmenas y por eso en la mayoría de las veces fuera de cualquier control.
Esta gran familia o superorganismo no puede estudiarse por las características aisladas e individuales de uno de sus miembros, mas bien por las características que la misma en su conjunto refleja. Por lo tanto si los portadores de las características genéticas son las REINAS y los ZÁNGANOS juzgaríamos a la reina no por las características individuales de ella, sino por las características de la super-familia o super-organismo que esta compone y de los ZÁNGANOS sabemos que son medio hermanos de la reina.
Además de lo complicado que resulta, por las razones antedichas, la aplicación de las leyes de la genética a la apicultura, tenemos los inconvenientes derivados de la partenogénesis en las reinas; el acoplamiento múltiple; acoplamiento con zánganos del origen desconocido; el hecho de que cada zángano muere en el acto del acoplamiento y por lo tanto no se puede utilizar para otros acoplamientos como es posible en otros ámbitos productivos. Y no olvidemos recordar que la reina se aparea muy joven y por única vez, después por el resto de su vida utiliza los espermas acumulados en la espermateca.(*) Aunque la abeja está conforme a los leyes universales válidas de la genéticas según lo establecido por MENDEL, sin embargo manifiesta excepciones y particularidades de vital importancia. PARTENOGÉNESIS (MITOS SOBRE EL ZÁNGANO)
La partenogénesis (del idioma griego παρθένος parthenos = virgen + γένεσις genesis = generación) es una forma de reproducción asexual por hembras no fecundadas , que se da con cierta frecuencia en insectos, anfibios y reptiles . Fue Jan Dzierzon quien primero descubrió la partenogénesis en los zánganos. Puede también considerarse como reproducción asexual o como sexual mono-gamética, puesto que interviene en ella una célula sexual o gameto
Como se sabe, las hembras de las abejas -reinas y obreras- nacen de un huevo, que es la unión de un óvulo de la reina con un espermatozoide de uno de los 10 a 15 zánganos que se aparearon con esa reina. En tanto que los zánganos nacen de los óvulos de la reina, esto significa que la reina produce zánganos sin necesidad de aparearse. Esta particularidad del sistema reproductivo de las abejas se denominó PARTENOGENESIS en 1845 cuando Dzierzon describe por primera vez el fenómeno. En 1986 el brillante apicultor uruguayo Samuel de León del Río, lo rebautizó como TELIGENESIS, en su muy interesante libro "He aquí las abejas" (Editorial Hemisferio Sur), y nos ayudó a comprender que los zánganos de una colmena son medios hermanos de la reina que puso el óvulo de donde nacieron, y no sus hijos como la teoría de la partenogénesis establecía.
Existen muchos mitos acerca del zángano. Siempre se afirmó que el zángano era hijo de la reina, cuando la realidad es que genéticamente es hermano de la reina, pues parte de una unidad biológica a la que mal se ha hecho en denominar huevo, ya que para que fuese un huevo debería contener la información genética de dos progenitores y en realidad es un óvulo formado a partir de una meiosis de las células de la reina.
Por esa razón es incorrecto decir que el zángano nace de un huevo infecundo porque en realidad ni es un huevo ni es infecundo. Podríamos llamarla MADRE HERMANA Para poder entender esto, puede citarse el caso de la mosca de la fruta o Ceratitis capitata. Esta mosca es combatida criando machos en cautiverio a los que se esteriliza en laboratorios para que luego al ser liberados fecunden a las hembras. Como los machos no tendrán semen al fecundar las hembras, éstas pondrán huevos que jamás incubarán por falta de uno de los gametos.
Si este mismo proceso se efectuara con las abejas y se esterilizaran los zánganos, ellas igualmente pondrían "huevos" que originarían machos.
Otro de los mitos existentes es el de afirmar que los zánganos serán todos iguales a su "madre hermana" cuando en realidad lo que sucede es lo contrario, pues la célula que origina al macho, ese óvulo hasta ahora llamado huevo infecundo, surge por una meiosis celular de la reina, es decir que de los 32 cromosomas apareados que forman el genoma de la reina sólo 16 pasarán al zángano.
EFECTOS DE LA PARTENOGÉNESIS
Aparte del hecho de que tenemos en el mejor de los casos, solamente un control limitado del acoplamiento de las reinas, es la partenogénesis el verdadero punto crítico en la crianza de la abeja. La partenogénesis anula, no solamente los procesos de crianza normales, también rompe todas nuestras nociones e hipótesis preconcebidas referentes a la herencia. Debido a la partenogénesis el zángano no tiene ningún padre sino solamente una madre “Madre Hermana”. Por otra parte él pierde su vida en el acto del acoplamiento y así deja de estar de servicio para los propósitos posteriores de la crianza. Por lo tanto no hay posibilidad en la abeja de acoplamientos entre el PADRE E HIJA, MADRE E HIJO (*), o HERMANO Y HERMANA. De máxima puede haber acoplamiento emparentado entre un HERMANASTRO Y UNA HERMANASTRA.
La situación es mas complicada por el hecho que, debido a la partenogénesis, millones de espermatozoides producidos por un zángano son todos absolutamente idénticos desde el punto de vista genético.
Un resultado de esta uniformidad de genes en el zángano significa que hay una mayor estabilidad en la herencia de la abeja que en otras formas de vida. Otra consecuencia de esta uniformidad es que la abeja es más susceptible a la endogamia. Es verdad que el acoplamiento múltiple actúa como contrapeso a esto, pero solamente parcialmente. En nuestro cruce obtenemos una segregación en el descendiente femenino en la F1, en los zánganos solamente en la F2, pero aquí no hay después el mismo patrón que en otros tipos de crianzas donde no hay partenogénesis. Cruzando a individuos de la F1 entre sí mismos, MENDEL podía obtener la segregación clásica en la F2, (Fenotipo 3:1) de la cual se presentan las nuevas combinaciones de los genes que entonces se transmiten en una línea directa. Tales nuevas combinaciones son posibles en el caso de la abeja, pero como se ha observado ya, solamente de una manera del cruce giratorio debido a el hecho de la partenogénesis.
El zángano no tiene ningún hijo, solamente nietos; él no tiene padre tiene solo madre y sus factores hereditarios correspondan a los de sus abuelos. Por lo tanto en un primer cruzamiento no tenemos ningún zángano de F 1, ellos aparecen solamente en la generación subsiguiente.
En las hembras fértiles se puede producir la hibridación y la segregación de genes en el primer cruzamiento (F1) pero no así en el zángano que es un mero multiplicador y transmisor de los gametos femeninos. En el zángano se produce la segregación recién en el segundo cruce o (F2)
Los zánganos nacen de un óvulo y tienen la mitad de cromosomas 16 y sus espermas tienen también 16 cromosomas por lo tanto repite las características genéticas del óvulo que lo engendró.
Las abejas al igual que las reinas tienen dos GAMETOS y por eso pueden ser “puras de una raza” o “mestizas”. En casos excepcionales desovan!. Los zánganos sólo tienen un GAMETO, nacen de un óvulo no fecundado por eso nacen PUROS DE ALGUNA RAZA ya que para el mestizaje habría necesidad de dos GAMETOS. Puede haber algún intercambio de genes por (“CROSSING-OVER”) o Sobrecruzamiento.
La espermateca de la REINA (local donde se quedan depositados vivos los espermatozoides de los zánganos) no tiene evidentemente, por lo tanto, la mínima influencia sobre su prole de zánganos (Ha no ser en sus hijas y nietas reinas). Sabedor de eso el apicultor o Genetista podrá programar nacimientos de zánganos aunque sus REINAS se hubieran fecundado con machos de otras razas. (*). Por eso una reina ITALIANA PURA aún fecundada por ZÁNGANOS AFRICANOS generará solamente ZÁNGANOS ITALIANOS. Si la reina fuera híbrida irá a producir simultáneamente dos razas diferentes de zánganos. Como ejemplo una Reina ITALIANA AFRICANIZADA irá a dar origen a la mitad de zánganos ITALIANOS (derivados del GAMETO “A )”y la otra mitad será de AFRICANOS (oriundos del GAMETO “B”). En mestizas es muy común encontrar un grupo de zánganos bien amarillos y otros bien oscuros
Por las Leyes de MENDEL son fácilmente recuperadas las RAZAS puras de entre las cruzas (mestizos). Esto se está haciendo hace más de 70 años en los Centros de Genética y sin que hayan ocurrido mayores complicaciones. Sobreestimar el SOBRECRUZAMENTO no explicaría porque después de tantos años de mestizajes en poblaciones cerrados y en constantes depuraciones no se quedó todo emparentado a tal punto que inviabilizase los desoves de aquellas reinas.
KK x K - KK x I - KI x K - KI x I
KK KK - KI KI - KK Ik - KI II
Primero Segundo Tercero Cuarto
En
el Gráfico de la izquierda (primero) todas las posibles PRINCESAS hijas
serán puras de la RAZA CÁRNICA (“KK”). En el segundo una CARNICA PURA
cruzada con un zángano ITALIANO da en F1 los mestizos CÁRNICOS
ITALIANIZADOS En la F2 cruzando el Híbrido KI con un zángano K (tercero) dará hijas de de la RAZA CÁRNICA Y otra vez un mestizo pero ahora el gameto del zángano esta del lado femenino IK dando un mestizo Italiano Carnicizado
En el de la derecha (cuarto)aparecen las ITALIANAS puras (“II”) y otra vez el híbrido invertido explicando como el gameto del zángano tiene segregación en la F2. Nadie puede negar esta Ley básica de MENDEL de que de los mestizos apareados reaparecen inmediatamente las RAZAS puras que les dieron el origen.
Es evidente que en BRASIL las APIS MELLÍFICA son africanas puras porque no hay abejas, reinas y zánganos europeos en los enjambres y ni los respectivos híbridos lo que sería una inevitabilidad científica; también tampoco existe la menor posibilidad aún usando las técnicas de la Inseminación Artificial de rescatar las razas antiguas pre-existentes a las africanas A. M. SCUTELLATA. De esto sólo puede ser explicada una conclusión lógica: las europeas y sus mestizas - las africanizadas - ya no existen y lo que hoy son, sí, solamente africanas puras (“AA”) cómo lo confirmó la cuenta de genes. Claudio Mikos
Es verdad que existe un concepto adicional: el del PEDIGRI. Las hijas del primer Gráfico podrían ser Pedigris si aquella MATRIZ (“KK – K”) tuviera un CERTIFICADO DE ORIGEN. En el caso de las CÁRNICAS (“KK”) y de las ITALIANAS (“II”) del Gráfico de la derecha a pesar de que sean tenidas cómo puras evidentemente no podrían ser calificadas como Pedigris. En el comercio de material genético como lo del ganado es usado el término “PO” (“PURO DE ORIGEN”) lo cual es cotizado por elevados precios. En el caso de las abejas es poco usual probablemente debido al hecho de que actualmente es muy difícil encontrar ejemplares sin que hayan sido mezclados en el pasado con otras razas. Hay además el detalle de que las reinas se aparean lejos de las colmenas y por eso en la mayoría de las veces fuera de cualquier control.
Como curiosidad por veces hay –
como ejemplo – mujeres negras con la piel totalmente negra a punto de
que nadie pueda negar que ellas sean puras de la raza negra, sin embargo
sus ojos son verde-claros! Las Leyes de MENDEL explican que entre los
apareamientos de individuos mestizos ellas recuperaron su raza negra
original y nuestros descubrimientos en el área de la genética explican
que en el final se quedaron con algunos genes de los blancos. Obviamente
no se podría decir que ellas sean “europeizadas” sólo por este detalle
como lo hace la élite apícola nacional al afirmar que las abejas del
BRASIL son “africanizadas”: están negando las evidencias de que son
africanas puras. Claudio Mikos
TENDENCIA AFRICANIZANTE
Para simplificar los Gráficos hicimos de cuenta que los zánganos eran sólo de una raza: o CÁRNICOS o AFRICANOS.
(“ F-0 “): Como se observa en el Gráfico de la izquierda el enjambre es CÁRNICO totalmente puro. Todas las abejas (“KK”) y las hijas PRINCESAS (“KK”) nacen puras de la raza CÁRNICA. Igualmente todos los zánganos allí creados son CÁRNICOS (“K”). Esta reina normalmente es importada o inseminada.
(“ F-1 ”): La colonia “F0” genera una hija de la “F2” (recuadro del centro). Está encabezada por una reina CÁRNICA PURA, pero que fue fecundada por zánganos AFRICANOS (“A”= a las A. M. SCUTELLATA). Aquí todas las futuras PRINCESAS y las abejas son y serán africanizadas, mestizas: CÁRNICAS AFRICANIZADAS (“KA”). Los zánganos aún son de la raza CÁRNICA (“K”). En este cambio de REINA MAESTRA ya está sufriendo la primera africanización porque la fecundación fue al natural - al aire libre - y con presencia de abejas africanas conjuntamente con sus zánganos en la región. La mansedumbre se mantiene prácticamente inalterada (“KA”): aún no nacen las obreras africanas puras (“AA”). Los GENES de las europeas en el aspecto de la mansedumbre son DOMINANTES en relación a los que responden por la agresividad de las africanas – RECESIVOS.
(“ F-2 “): El enjambre ya está encabezado por una REINA mestiza (nieta): CÁRNICA AFRICANIZADA (“KA”); es decir; a sólo un paso de la africanización total. En esta solamente el GAMETO “A ” de la reina es europeo, EL “B ” es africano y fue fecundada por zánganos africanos (“A” ). Como se ve en el recuadro de la derecha mitad de las obreras nacen mestizas igual a la madre (africanizadas: “KA”) y la otra son africanas puras (“AA”). En esta familia mitad de los zánganos será de la raza CÁRNICA (“K”) mientras la otra será AFRICANA (“A )”. En esta familia ya hay AGRESIVIDAD porque un 50% de las obreras nacen africanas puras “AA”. Solamente un 50% de las obreras aún tienen un GAMETO CÁRNICO (“KA”). Claudio Mikos
Hay fundamentalmente dos hechos que favorecen a las reinas africanas para tener ventajas en la propagación de la raza
1-- Las PRINCESAS africanas puras nacen un dia antes que las mestizas y cerca de dos antes que las europeas puras;
Como en los cambios y en las enjambrazones la que nace primero normalmente elimina las nuevas todavía en sus celdillas, las probabilidades de una nueva mestiza venir a defenderse son prácticamente nulas;
2- El Otro el es hecho de que algunas TRIBOS dejan nacer varias nuevas PRINCESAS e impiden que luchen entre si manteniéndolas separadas. Allí la primera que consiga iniciar la postura será la madre. En esta situación las africanas puras también tienen ventaja.
CRUZAS SIMPLES (Mestizajes)
Se dice mucho sobre la creación de híbridos entre las abejas pero realmente las hibridaciones se producen al cruzar dos especies distintas como ocurre en la MULA hija de un caballo y el asno o burro. La mula tiene en denominado vigor híbrido pero es esteril, lo mismo ocurre con otro tipo de híbridos pero en las abejas lo que se cruza son subespecies o razas a veces hasta tribus diferentes de una misma raza y no híbridos como se quiere hacer creer. La cruza entre distintas especies de Apis, como la mellifica, dorsata, florea, cerana, etc no es viable.
Muy frecuentes son los mestizajes simples: el apicultor compra la REINA pura de una raza ya apareada con zánganos de la misma. Después crea las PRINCESAS y estas se aparean con los zánganos de otra raza de abejas existentes en la región. Es el caso típico de los criadores que importan las REINAS de Europa y las PRINCESAS-hijas se aparean con los zánganos africanos de las A. M. SCUTELLATA. Si el lado europeo tiene buenas características y también el africano resulta en un mestizo dócil, prolífico y altamente productivo.
PEDIGRÍ EN LAS ABEJAS
El máximo de expresión de una raza es un PEDIGRI.
El estudio del pedigrí es un concepto utilizado desde los tiempos remotos como método de pureza de raza, es un término que se refiere a la GENEALOGIA del animal, a su ascendencia biológica
La certificación de pedigrí reconoce a un animal como perteneciente a una RAZA que ha sido reconocida por su belleza, originalidad, performance, etc. y/o alguna característica deseable.
Un formato pedigrí es generalmente un diagrama estandardizado, demostrando simplemente una línea del padre y una de la madre a unos o más descendiente.
Debido a QUE los zánganos provienen de un óvulo, los pedigríes de la abeja son diferentes. (*)
(*)”Normalmente entre animales cada individuo tiene dos padres, cuatro abuelos, ocho bis-abuelos, dieciséis tártara-abuelos, etc., el número siempre doble de sus antepasados inmediatos. En el caso de la abeja el zángano tiene solo su madre, dos abuelos, tres bis-abuelos y solamente cinco tártara-abuelos. Las reinas y las obreras tienen dos padres, pero solamente tres abuelos, cinco bis-abuelos y ocho tártara-abuelos, es decir, solamente mitad del número según la regla general. Pero el zángano no tiene exactamente la mitad del número según la regla. Está claro de esto que el pedigrí de la abeja diferencia esencialmente al de otras formas de seres vivos. Es no sólo el pedigrí de ambos sexos diferente en lo que concierne al número de antepasados, sino que el zángano no tiene ni el padre ni hijos; él tiene solamente un abuelo y nietos. Además de esto el pedigrí de la abeja llega a ser aún más complicado debido a el acoplamiento múltiple” Hermano Adan.
Para discutir correctamente un pedigrí de la abeja, dos términos deben ser definidos. Éstas son: Gameto y Segregación.
GAMETO:
En Biología, los gametos (del griego Gameto, cónyuge) son cada una de las células sexuales ... Los órganos que producen los gametos se llaman gónadas .La hembra produce los óvulos y el macho los espermatozoides mitad de los cromosomas necesitados para producir a una abeja obrera a una reina.
SEGREGACIÓN:
Una de las maneras en que la meiosis produce variación genética, es a través de las diferentes formas que los cromosomas maternal y paternal son combinados en las células hijas. Sucede al clasificar al azar de los cromosomas apareados para producir gametos. En la mayoría de los animales, la segregación ocurre en los ovarios y en los testículos.
En las abejas, la segregación ocurre solamente en los ovarios de las reinas.
Por lo tanto, en las abejas, todos los nuevos gametos se originan de una reina. Decimos "nuevos" gametos porque los zánganos propagan solamente gametos existentes.
Los zánganos entonces tienen dos funciones reproductivas:
-Primero: Convierten y extienden el gameto femenino de la reina en cerca de 10 millones de gametos masculinos idénticos (espermas).
-En segundo lugar, sirven como vehículo para mover los gametos propagados a la reina (en el acto del acoplamiento).
En la reproducción de la abeja, entonces, la progenie femenina recibe un gameto de la reina que produjo el huevo y el otro gameto de otra reina (vía la conversión del zángano del gameto a una célula de la esperma).
Así, un pedigrí de la abeja contiene solamente a hembras y solamente las hembras (o las reinas) necesitan ser registradas. (Juan R. Harbo y Thomas E. Rinderer
COMPORTAMIENTO HIGIÉNICO
El comportamiento higiénico es probablemente el logro más acertado en la cría de abejas. Se estudia y se demuestra muy bien que es eficaz en el control de ser eficaz en Cría yesificada, Loque Americana, y Varroa
 El
comportamiento higiénico es controlado por dos genes. RECESIVOS Un gen
permite que la obrera detecte y destape una célula que contenga la cría
enferma. El otro gen hace que las obreras quitan la cría y la desechen.
Es posible que una colmena contenga uno de estos genes y no el otro, en
este caso la colmena no será higiénica. Es también posible que diversas
abejas individuales tengan uno, pero no el otro gen.
El
comportamiento higiénico es controlado por dos genes. RECESIVOS Un gen
permite que la obrera detecte y destape una célula que contenga la cría
enferma. El otro gen hace que las obreras quitan la cría y la desechen.
Es posible que una colmena contenga uno de estos genes y no el otro, en
este caso la colmena no será higiénica. Es también posible que diversas
abejas individuales tengan uno, pero no el otro gen.
Para que un rasgo recesivo sea expresado, una obrera necesita ser homocigótica para el gen. Homocigótico significa que consigue el mismo alelo de la madre y del padre. Heterocigótico significa que la abeja tiene uno de los alelos y así que es un portador, pero el rasgo no se expresa.
En este ejemplo estamos comenzando con una reina homocigótica para los rasgos higiénicos y acoplarla a los abejones no higiénicos. El descendiente no expresará el rasgo higiénico, sino que serán heterocigóticos pero serán portadores del rasgo. Es importante recordar que cuando usted se está ocupando de rasgos recesivos, estos no se evidencian en la primer generación F1 sino mas tarde en la segunda generación la F2.
Después de algunas generaciones de seleccionar las colonias que expresan el rasgo, puede llegar a ser fijo en la población. Entonces todas las abejas en esa población expresarán el rasgo. Hemos alcanzado este punto con la inseminación artificial y poblaciones cerradas.
¿CÓMO SE DETERMINA EL COMPORTAMIENTO HIGIÉNICO? Hay varios métodos para determinar el índice de comportamiento higienico. El mas efectivo es por medio del nitrógeno líquido. Se congela una porción de un panal con cría operculada y luego se vuelve a colocar al mismo en la colonia para luego determinar el índice de celdillas vacíadas por las abejas.
También se puede cortar un trozo de panal y congelar en una nevera y luego injertar en el panal dentro de la colonia.
Y por último: el método mas práctico para un principiante consiste en utilizar un alfiler para matar la cría y después que esta es retirada por la abeja, evaluar la incidencia del comportamiento higiénico.
1) marcar una celda directamente sobre tres grupos de siete celdas nuevamente selladas. Utilice una pintura que seque rápido (e.g. papel líquido). También marque el cabezal del cuadro.
2) Mate a las veintiún larvas pinchándolas con un alfiler a través de los opérculos. Utilice el mismo agujero para pinchar la larva varias veces a diversos ángulos.
3) Veinticuatro horas más adelante cuente cuántas celdas se destapan y se limpian totalmente hacia fuera. Se considera a una colonia que retiró 19 larvas como 90% higiénica.
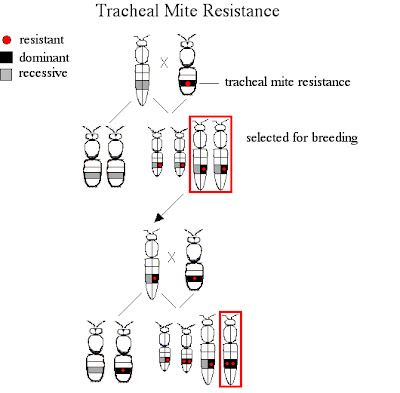
RESISTENCIA A LOS ÁCAROS TRAQUEALES Las abejas utilizan sus piernas medias para RETIRAR los ácaros lejos de su abertura traqueal. También se encuentra que el rasgo es controlado por los genes DOMINANTES. No se ha determinado si hay más de un gen implicado. En este ejemplo, asumiremos que hay apenas un gene que lo controla. Aquí diremos que estamos comenzando con un solo abejón que lleve el gene resistente.
ABEJAS SMR La “Reproducción Suprimida del ácaro” (SMR) es la última técnica que se está estudiando. Hay gran expectativa de solucionar el gran problema que causa la varroa con esta técnica, pero es demasiado temprano para decir exactamente cómo y porqué actúa. Hay algo en esas abejas que llevan el rasgo que inhibe en los ácaros varroa su reproducción normal. Algunos de los ácaros no ponen ningún huevo. Otros ponen los huevos demasiado tarde en el ciclo . Y otros colocan solamente un huevo masculino. Otros son retirados de entre el capullo de la larva de la abeja y la pared de célula antes de que puedan poner huevos. Si todas formas del rasgo son el efecto de un factor o de varios siguen siendo una incógnita. También no sabemos cuántos genes están implicados. Afortunadamente, no es necesario saber todos estos detalles para seleccionar y para utilizar el rasgo.
 El Dr. Harbo y el Dr.
Harris han criado selectivamente las abejas para este rasgo al punto en
que pocos y casi ningún ácaros se reproducen normalmente. Cruzando estas
abejas innatas con las abejas de SMR encontraron que los efectos eran
intermedios entre los dos tipos. Esto indica que el rasgo no es
controlado ni por los genes dominantes ni por los recesivos, en
realidad es ADITIVO. Esto significa simplemente que cuanto más de estos
genes están presentes, mas se expresa el rasgo.
El Dr. Harbo y el Dr.
Harris han criado selectivamente las abejas para este rasgo al punto en
que pocos y casi ningún ácaros se reproducen normalmente. Cruzando estas
abejas innatas con las abejas de SMR encontraron que los efectos eran
intermedios entre los dos tipos. Esto indica que el rasgo no es
controlado ni por los genes dominantes ni por los recesivos, en
realidad es ADITIVO. Esto significa simplemente que cuanto más de estos
genes están presentes, mas se expresa el rasgo.
EL ADN MITOCONDRIAL Tradicionalmente,
la taxonomía intraespecífica de la abeja melífera, Apis mellífera, se
ha basado en la morfología. En el presente, están reconocidas 26
subespecies de A. mellifera, teniendo en cuenta suscaracteres morfométricos (RUTTNER, 1988, 1992; SHEPPARD et al., 1997).
En fechas más recientes, los instrumentos genéticos, principalmente el análisis de la secuencia ADN y la electroforesis con alosima, se aplicaron al estudio de la diversidad de la abeja melífera. El ADN mitocondrial (mtADN) posee ciertas propiedades que le convierten en instrumento favorito en la sistemática y la biología de las poblaciones. En general, es heredado por vía materna, sin recombinación. Por tanto, permite detectar con precisión los haplotipos extraños dentro de las poblaciones. Sólo la herencia materna del mtADN quedó comprobada en las abejas melíferas (MEUSEL y MORITZ, 1993) y, así, todas las obreras y todos los zánganos de una colonia comparten con la reina el mismo mtADN. Comisión Permanente de Biología Apícola ESTRUCTURA GENETICA DE LAS ABEJAS DE LA ISLA DE CRETA (GRECIA)P. HARIZANIS, Maria BOUGA Laboratorio de sericicultura – apicultura, Universidad agrícola de Atenas, 75, Iera Odos, 118 55, Atenas, GRECIA
Las mitocondrias son los pequeños organelos que se encuentran en cada célula viva. Las mitocondrias son esenciales para la producción de energía, están presentes en el citoplasma de las células y - lo que es relevante aquí - poseen su propio ADN. son los qué hacen posible la respiración. Liberan energía quemando azúcar con el oxigeno.
La cosa importante a saber sobre las mitocondrias es que se reproducen por separado del resto de la célula. Sucede que cuando ocurre el proceso de fecundación del óvulo por el espermatozoide, sólo el núcleo del espermatozoide logra penetrar el óvulo. En consecuencia, las mitocondrias (y su ADN mitocondrial) del nuevo ser engendrado provienen exclusivamente del óvulo, es decir de la madre. Cuando las células se dividen, las mitocondrias se dividen en el mismo tiempo. Contienen una cantidad pequeña de ADN, pero este ADN permanece separado del núcleo. Las mitocondrias están presentes en los óvulos cuando se forman. Pero cuando el esperma se une con el núcleo en la fertilización para crear una nueva combinación genética, las mitocondrias permanecen sin cambiar. Consiguen permanecer de generación en generación a través de los huevos sin su ADN, que es afectada siempre por los espermas. Son transmitidos solamente a través de sus madres, y su ADN en ellas cambia solamente muy lentamente por mutaciones ocasionales.
Una gran cantidad de mitocondria se encuentran en la cola del esperma, la que aporta la energía, cual un motor, que permite nadar al esperma hacia el óvulo. Sin embargo, cuando el esperma entra en el huevo durante la fertilización, la cola se cae, quitando las mitocondrias del padre
A partir del estudio del ADN mitocondrial se puede conocer el linaje de la abeja y diferenciar un tipo de abeja de otro. Es la forma en que se logran distinguir las abejas africanas de las europeas.
LAS MUTACIONES.- Las mutaciones son cambios que se producen en la duplicación de los genes.- Una vez mutado un gen a una nueva forma, la misma es estable y sin mayor tendencia a mutar.
Las mutaciones cromósicas causan cambios visibles en la estructura del cromosoma. Pueden ser:
1.- por delección: falta de un segmento del cromosoma
2.- por duplicación: cuando un segmento está representados dos veces
3.- por traslocación: cuando un segmento cromosómico puede ser traslocado sobre un nuevo cromosoma
4.-por inversión: cuando un segmento cromosómico se adhiere al otro extremo del mismo cromosoma.
a b c d e f normal
a b d e f supresión (segmento c)
a b c c d e f duplicación (segmento c)
a e d c b f inversión (segmento b-e)
Las mutaciones de genes suelen depender de errores en el apareamiento de las bases en el proceso de la duplicación. El D.N.A. modificado será transcrito y producirá A.R.N. m alterado, el cual será descifrado con un aminoácido diferente del normal. Lo anterior producirá proteínas alteradas.
Las mutaciones del gen pueden deberse a la acción de rayos X, rayos gamma, rayos cósmicos, rayos ultravioletas.
Algunos genes (puntos calientes) son más suceptibles que otros de las mutaciones.
A nivel molecular una mutación es una alteración de la secuencia de las bases púricas o pirimídicas en las cadenas del ácido nucleico.
Más de 30 mutaciones visibles específicas se han descrito en abejas, y un número éstos son mantenidos por los laboratorios de investigación. Generalmente, estas mutaciones producen un efecto llamativo, y a sus descubridores ha observado a la mayoría fácilmente. Muchas otras mutaciones pudieron ocurrir en las abejas que también hacen cambios sutiles todavía ser observadas. Las mutaciones sabidas afectan el color, la forma, y la presencia de ojos, el color y la pilosidad de cuerpos, la forma y tamaño de alas, y del comportamiento.
Probablemente debido a su aspecto distintivo, la mayoría de los mutantes de la abeja hasta el momento recogidos tenían variaciones en el color de ojos.
Además de contribuir al trabajo sobre la bioquímica del pigmento del ojo, los mutantes se han utilizado como herramientas para investigar una variedad de otros interrogantes. La resistencia al Loque americano, determinación de sexo, actividad de la polinización, tecnología de la fertilización, almacenaje de la esperma, dinámica de la población, longevidad, Debido a esta utilidad y de otros usos potenciales, es deseable que la comunidad científica mantenga un número de mutaciones. Las mutaciones nuevamente descubiertas pueden tener usos especiales en la ciencia; por lo tanto, los apicultores podrían ayudar informando sobre las nuevas mutaciones que observan a un laboratorio de investigación.
 La mayoría de las mutaciones son recesivas. Las
mutaciones, por lo tanto, son a menudo observadas primero en los
zánganos, porque los zánganos son haploides y no enmascaran genes
recesivos. Una mutación puede ocurrir en un solo zángano en una colonia o
en muchos zánganos.
La mayoría de las mutaciones son recesivas. Las
mutaciones, por lo tanto, son a menudo observadas primero en los
zánganos, porque los zánganos son haploides y no enmascaran genes
recesivos. Una mutación puede ocurrir en un solo zángano en una colonia o
en muchos zánganos.
Esta variación del color del ojo cambia de amarillo ligero en un cierto plazo a rojo oscuro
Un gen recesivo produce ojos blancos. Las abejas de ojos blancos se comportan normalmente pero demoran en tomar su primer vuelo y cuando salen de la colmena vuelan en círculos, ellas nunca vuelve a la colmena.
TENDENCIA AFRICANIZANTE
F0 F1 F2 F3
KK x K KK x A KA x A AA x A
KK KK KA KA KA AA AA AA
En
Sudamérica con la llegada de la abeja africana en muy poco tiempo esta
ganó terreno y las abejas locales desaparecieron dando lugar a las
africanas puras Apis Mellifica Scutellata . En el gráfico de arriba se
ve como partiendo de una reina CÁRNICA pura cuando esta es fecundada por
zánganos AFRICANOS en muy poco tiempo ya se tiene todo el colmenar de
la raza Africana pura A M Scutellata Claudio Mikos Para simplificar los Gráficos hicimos de cuenta que los zánganos eran sólo de una raza: o CÁRNICOS o AFRICANOS.
(“ F-0 “): Como se observa en el Gráfico de la izquierda el enjambre es CÁRNICO totalmente puro. Todas las abejas (“KK”) y las hijas PRINCESAS (“KK”) nacen puras de la raza CÁRNICA. Igualmente todos los zánganos allí creados son CÁRNICOS (“K”). Esta reina normalmente es importada o inseminada.
(“ F-1 ”): La colonia “F0” genera una hija de la “F2” (recuadro del centro). Está encabezada por una reina CÁRNICA PURA, pero que fue fecundada por zánganos AFRICANOS (“A”= a las A. M. SCUTELLATA). Aquí todas las futuras PRINCESAS y las abejas son y serán africanizadas, mestizas: CÁRNICAS AFRICANIZADAS (“KA”). Los zánganos aún son de la raza CÁRNICA (“K”). En este cambio de REINA MAESTRA ya está sufriendo la primera africanización porque la fecundación fue al natural - al aire libre - y con presencia de abejas africanas conjuntamente con sus zánganos en la región. La mansedumbre se mantiene prácticamente inalterada (“KA”): aún no nacen las obreras africanas puras (“AA”). Los GENES de las europeas en el aspecto de la mansedumbre son DOMINANTES en relación a los que responden por la agresividad de las africanas – RECESIVOS.
(“ F-2 “): El enjambre ya está encabezado por una REINA mestiza (nieta): CÁRNICA AFRICANIZADA (“KA”); es decir; a sólo un paso de la africanización total. En esta solamente el GAMETO “A ” de la reina es europeo, EL “B ” es africano y fue fecundada por zánganos africanos (“A” ). Como se ve en el recuadro de la derecha mitad de las obreras nacen mestizas igual a la madre (africanizadas: “KA”) y la otra son africanas puras (“AA”). En esta familia mitad de los zánganos será de la raza CÁRNICA (“K”) mientras la otra será AFRICANA (“A )”. En esta familia ya hay AGRESIVIDAD porque un 50% de las obreras nacen africanas puras “AA”. Solamente un 50% de las obreras aún tienen un GAMETO CÁRNICO (“KA”). Claudio Mikos
Hay fundamentalmente dos hechos que favorecen a las reinas africanas para tener ventajas en la propagación de la raza
1-- Las PRINCESAS africanas puras nacen un dia antes que las mestizas y cerca de dos antes que las europeas puras;
Como en los cambios y en las enjambrazones la que nace primero normalmente elimina las nuevas todavía en sus celdillas, las probabilidades de una nueva mestiza venir a defenderse son prácticamente nulas;
2- El Otro el es hecho de que algunas TRIBOS dejan nacer varias nuevas PRINCESAS e impiden que luchen entre si manteniéndolas separadas. Allí la primera que consiga iniciar la postura será la madre. En esta situación las africanas puras también tienen ventaja.
CRUZAS SIMPLES (Mestizajes)
Se dice mucho sobre la creación de híbridos entre las abejas pero realmente las hibridaciones se producen al cruzar dos especies distintas como ocurre en la MULA hija de un caballo y el asno o burro. La mula tiene en denominado vigor híbrido pero es esteril, lo mismo ocurre con otro tipo de híbridos pero en las abejas lo que se cruza son subespecies o razas a veces hasta tribus diferentes de una misma raza y no híbridos como se quiere hacer creer. La cruza entre distintas especies de Apis, como la mellifica, dorsata, florea, cerana, etc no es viable.
Muy frecuentes son los mestizajes simples: el apicultor compra la REINA pura de una raza ya apareada con zánganos de la misma. Después crea las PRINCESAS y estas se aparean con los zánganos de otra raza de abejas existentes en la región. Es el caso típico de los criadores que importan las REINAS de Europa y las PRINCESAS-hijas se aparean con los zánganos africanos de las A. M. SCUTELLATA. Si el lado europeo tiene buenas características y también el africano resulta en un mestizo dócil, prolífico y altamente productivo.
CC x C CC x A
CC CC CA CA
CC CC CA CA
El máximo de expresión de una raza es un PEDIGRI.
El estudio del pedigrí es un concepto utilizado desde los tiempos remotos como método de pureza de raza, es un término que se refiere a la GENEALOGIA del animal, a su ascendencia biológica
La certificación de pedigrí reconoce a un animal como perteneciente a una RAZA que ha sido reconocida por su belleza, originalidad, performance, etc. y/o alguna característica deseable.
Un formato pedigrí es generalmente un diagrama estandardizado, demostrando simplemente una línea del padre y una de la madre a unos o más descendiente.
Debido a QUE los zánganos provienen de un óvulo, los pedigríes de la abeja son diferentes. (*)
(*)”Normalmente entre animales cada individuo tiene dos padres, cuatro abuelos, ocho bis-abuelos, dieciséis tártara-abuelos, etc., el número siempre doble de sus antepasados inmediatos. En el caso de la abeja el zángano tiene solo su madre, dos abuelos, tres bis-abuelos y solamente cinco tártara-abuelos. Las reinas y las obreras tienen dos padres, pero solamente tres abuelos, cinco bis-abuelos y ocho tártara-abuelos, es decir, solamente mitad del número según la regla general. Pero el zángano no tiene exactamente la mitad del número según la regla. Está claro de esto que el pedigrí de la abeja diferencia esencialmente al de otras formas de seres vivos. Es no sólo el pedigrí de ambos sexos diferente en lo que concierne al número de antepasados, sino que el zángano no tiene ni el padre ni hijos; él tiene solamente un abuelo y nietos. Además de esto el pedigrí de la abeja llega a ser aún más complicado debido a el acoplamiento múltiple” Hermano Adan.
Para discutir correctamente un pedigrí de la abeja, dos términos deben ser definidos. Éstas son: Gameto y Segregación.
GAMETO:
En Biología, los gametos (del griego Gameto, cónyuge) son cada una de las células sexuales ... Los órganos que producen los gametos se llaman gónadas .La hembra produce los óvulos y el macho los espermatozoides mitad de los cromosomas necesitados para producir a una abeja obrera a una reina.
SEGREGACIÓN:
Una de las maneras en que la meiosis produce variación genética, es a través de las diferentes formas que los cromosomas maternal y paternal son combinados en las células hijas. Sucede al clasificar al azar de los cromosomas apareados para producir gametos. En la mayoría de los animales, la segregación ocurre en los ovarios y en los testículos.
En las abejas, la segregación ocurre solamente en los ovarios de las reinas.
Por lo tanto, en las abejas, todos los nuevos gametos se originan de una reina. Decimos "nuevos" gametos porque los zánganos propagan solamente gametos existentes.
Los zánganos entonces tienen dos funciones reproductivas:
-Primero: Convierten y extienden el gameto femenino de la reina en cerca de 10 millones de gametos masculinos idénticos (espermas).
-En segundo lugar, sirven como vehículo para mover los gametos propagados a la reina (en el acto del acoplamiento).
En la reproducción de la abeja, entonces, la progenie femenina recibe un gameto de la reina que produjo el huevo y el otro gameto de otra reina (vía la conversión del zángano del gameto a una célula de la esperma).
Así, un pedigrí de la abeja contiene solamente a hembras y solamente las hembras (o las reinas) necesitan ser registradas. (Juan R. Harbo y Thomas E. Rinderer
COMPORTAMIENTO HIGIÉNICO
El comportamiento higiénico es probablemente el logro más acertado en la cría de abejas. Se estudia y se demuestra muy bien que es eficaz en el control de ser eficaz en Cría yesificada, Loque Americana, y Varroa
Para que un rasgo recesivo sea expresado, una obrera necesita ser homocigótica para el gen. Homocigótico significa que consigue el mismo alelo de la madre y del padre. Heterocigótico significa que la abeja tiene uno de los alelos y así que es un portador, pero el rasgo no se expresa.
En este ejemplo estamos comenzando con una reina homocigótica para los rasgos higiénicos y acoplarla a los abejones no higiénicos. El descendiente no expresará el rasgo higiénico, sino que serán heterocigóticos pero serán portadores del rasgo. Es importante recordar que cuando usted se está ocupando de rasgos recesivos, estos no se evidencian en la primer generación F1 sino mas tarde en la segunda generación la F2.
Después de algunas generaciones de seleccionar las colonias que expresan el rasgo, puede llegar a ser fijo en la población. Entonces todas las abejas en esa población expresarán el rasgo. Hemos alcanzado este punto con la inseminación artificial y poblaciones cerradas.
¿CÓMO SE DETERMINA EL COMPORTAMIENTO HIGIÉNICO? Hay varios métodos para determinar el índice de comportamiento higienico. El mas efectivo es por medio del nitrógeno líquido. Se congela una porción de un panal con cría operculada y luego se vuelve a colocar al mismo en la colonia para luego determinar el índice de celdillas vacíadas por las abejas.
También se puede cortar un trozo de panal y congelar en una nevera y luego injertar en el panal dentro de la colonia.
Y por último: el método mas práctico para un principiante consiste en utilizar un alfiler para matar la cría y después que esta es retirada por la abeja, evaluar la incidencia del comportamiento higiénico.
1) marcar una celda directamente sobre tres grupos de siete celdas nuevamente selladas. Utilice una pintura que seque rápido (e.g. papel líquido). También marque el cabezal del cuadro.
2) Mate a las veintiún larvas pinchándolas con un alfiler a través de los opérculos. Utilice el mismo agujero para pinchar la larva varias veces a diversos ángulos.
3) Veinticuatro horas más adelante cuente cuántas celdas se destapan y se limpian totalmente hacia fuera. Se considera a una colonia que retiró 19 larvas como 90% higiénica.
RESISTENCIA A LOS ÁCAROS TRAQUEALES Las abejas utilizan sus piernas medias para RETIRAR los ácaros lejos de su abertura traqueal. También se encuentra que el rasgo es controlado por los genes DOMINANTES. No se ha determinado si hay más de un gen implicado. En este ejemplo, asumiremos que hay apenas un gene que lo controla. Aquí diremos que estamos comenzando con un solo abejón que lleve el gene resistente.
Los
rasgos dominantes son más fáciles de establecer en la población porque
la primera generación expresará el rasgo (F1). El rasgo será expresado
igualmente por las abejas que llevan el gen para el rasgo en uno o ambos
de sus cromosomas . Usted preferiría criar solamente reinas
homocigóticas para el rasgo. Pero no es fácil identificar los
homocigotas de los heterocigotas. Por esta razón es mas difícil fijar un
rasgo dominante en la población que un recesivo.
ABEJAS SMR La “Reproducción Suprimida del ácaro” (SMR) es la última técnica que se está estudiando. Hay gran expectativa de solucionar el gran problema que causa la varroa con esta técnica, pero es demasiado temprano para decir exactamente cómo y porqué actúa. Hay algo en esas abejas que llevan el rasgo que inhibe en los ácaros varroa su reproducción normal. Algunos de los ácaros no ponen ningún huevo. Otros ponen los huevos demasiado tarde en el ciclo . Y otros colocan solamente un huevo masculino. Otros son retirados de entre el capullo de la larva de la abeja y la pared de célula antes de que puedan poner huevos. Si todas formas del rasgo son el efecto de un factor o de varios siguen siendo una incógnita. También no sabemos cuántos genes están implicados. Afortunadamente, no es necesario saber todos estos detalles para seleccionar y para utilizar el rasgo.
El Dr. Harbo y el Dr.
Harris han criado selectivamente las abejas para este rasgo al punto en
que pocos y casi ningún ácaros se reproducen normalmente. Cruzando estas
abejas innatas con las abejas de SMR encontraron que los efectos eran
intermedios entre los dos tipos. Esto indica que el rasgo no es
controlado ni por los genes dominantes ni por los recesivos, en
realidad es ADITIVO. Esto significa simplemente que cuanto más de estos
genes están presentes, mas se expresa el rasgo.En fechas más recientes, los instrumentos genéticos, principalmente el análisis de la secuencia ADN y la electroforesis con alosima, se aplicaron al estudio de la diversidad de la abeja melífera. El ADN mitocondrial (mtADN) posee ciertas propiedades que le convierten en instrumento favorito en la sistemática y la biología de las poblaciones. En general, es heredado por vía materna, sin recombinación. Por tanto, permite detectar con precisión los haplotipos extraños dentro de las poblaciones. Sólo la herencia materna del mtADN quedó comprobada en las abejas melíferas (MEUSEL y MORITZ, 1993) y, así, todas las obreras y todos los zánganos de una colonia comparten con la reina el mismo mtADN. Comisión Permanente de Biología Apícola ESTRUCTURA GENETICA DE LAS ABEJAS DE LA ISLA DE CRETA (GRECIA)P. HARIZANIS, Maria BOUGA Laboratorio de sericicultura – apicultura, Universidad agrícola de Atenas, 75, Iera Odos, 118 55, Atenas, GRECIA
Las mitocondrias son los pequeños organelos que se encuentran en cada célula viva. Las mitocondrias son esenciales para la producción de energía, están presentes en el citoplasma de las células y - lo que es relevante aquí - poseen su propio ADN. son los qué hacen posible la respiración. Liberan energía quemando azúcar con el oxigeno.
La cosa importante a saber sobre las mitocondrias es que se reproducen por separado del resto de la célula. Sucede que cuando ocurre el proceso de fecundación del óvulo por el espermatozoide, sólo el núcleo del espermatozoide logra penetrar el óvulo. En consecuencia, las mitocondrias (y su ADN mitocondrial) del nuevo ser engendrado provienen exclusivamente del óvulo, es decir de la madre. Cuando las células se dividen, las mitocondrias se dividen en el mismo tiempo. Contienen una cantidad pequeña de ADN, pero este ADN permanece separado del núcleo. Las mitocondrias están presentes en los óvulos cuando se forman. Pero cuando el esperma se une con el núcleo en la fertilización para crear una nueva combinación genética, las mitocondrias permanecen sin cambiar. Consiguen permanecer de generación en generación a través de los huevos sin su ADN, que es afectada siempre por los espermas. Son transmitidos solamente a través de sus madres, y su ADN en ellas cambia solamente muy lentamente por mutaciones ocasionales.
Una gran cantidad de mitocondria se encuentran en la cola del esperma, la que aporta la energía, cual un motor, que permite nadar al esperma hacia el óvulo. Sin embargo, cuando el esperma entra en el huevo durante la fertilización, la cola se cae, quitando las mitocondrias del padre
A partir del estudio del ADN mitocondrial se puede conocer el linaje de la abeja y diferenciar un tipo de abeja de otro. Es la forma en que se logran distinguir las abejas africanas de las europeas.
LAS MUTACIONES.- Las mutaciones son cambios que se producen en la duplicación de los genes.- Una vez mutado un gen a una nueva forma, la misma es estable y sin mayor tendencia a mutar.
Las mutaciones cromósicas causan cambios visibles en la estructura del cromosoma. Pueden ser:
1.- por delección: falta de un segmento del cromosoma
2.- por duplicación: cuando un segmento está representados dos veces
3.- por traslocación: cuando un segmento cromosómico puede ser traslocado sobre un nuevo cromosoma
4.-por inversión: cuando un segmento cromosómico se adhiere al otro extremo del mismo cromosoma.
a b c d e f normal
a b d e f supresión (segmento c)
a b c c d e f duplicación (segmento c)
a e d c b f inversión (segmento b-e)
Las mutaciones de genes suelen depender de errores en el apareamiento de las bases en el proceso de la duplicación. El D.N.A. modificado será transcrito y producirá A.R.N. m alterado, el cual será descifrado con un aminoácido diferente del normal. Lo anterior producirá proteínas alteradas.
Las mutaciones del gen pueden deberse a la acción de rayos X, rayos gamma, rayos cósmicos, rayos ultravioletas.
Algunos genes (puntos calientes) son más suceptibles que otros de las mutaciones.
A nivel molecular una mutación es una alteración de la secuencia de las bases púricas o pirimídicas en las cadenas del ácido nucleico.
Más de 30 mutaciones visibles específicas se han descrito en abejas, y un número éstos son mantenidos por los laboratorios de investigación. Generalmente, estas mutaciones producen un efecto llamativo, y a sus descubridores ha observado a la mayoría fácilmente. Muchas otras mutaciones pudieron ocurrir en las abejas que también hacen cambios sutiles todavía ser observadas. Las mutaciones sabidas afectan el color, la forma, y la presencia de ojos, el color y la pilosidad de cuerpos, la forma y tamaño de alas, y del comportamiento.
Probablemente debido a su aspecto distintivo, la mayoría de los mutantes de la abeja hasta el momento recogidos tenían variaciones en el color de ojos.
Además de contribuir al trabajo sobre la bioquímica del pigmento del ojo, los mutantes se han utilizado como herramientas para investigar una variedad de otros interrogantes. La resistencia al Loque americano, determinación de sexo, actividad de la polinización, tecnología de la fertilización, almacenaje de la esperma, dinámica de la población, longevidad, Debido a esta utilidad y de otros usos potenciales, es deseable que la comunidad científica mantenga un número de mutaciones. Las mutaciones nuevamente descubiertas pueden tener usos especiales en la ciencia; por lo tanto, los apicultores podrían ayudar informando sobre las nuevas mutaciones que observan a un laboratorio de investigación.
La mayoría de las mutaciones son recesivas. Las
mutaciones, por lo tanto, son a menudo observadas primero en los
zánganos, porque los zánganos son haploides y no enmascaran genes
recesivos. Una mutación puede ocurrir en un solo zángano en una colonia o
en muchos zánganos.Esta variación del color del ojo cambia de amarillo ligero en un cierto plazo a rojo oscuro
Un gen recesivo produce ojos blancos. Las abejas de ojos blancos se comportan normalmente pero demoran en tomar su primer vuelo y cuando salen de la colmena vuelan en círculos, ellas nunca vuelve a la colmena.
Si la madre y el padre contribuyen un gen blanco del ojo, las obreras y las reinas nacen con ojos blancos.

Combinaciones posibles de las abejas con ojos blancos.
No hay comentarios:
Publicar un comentario
Gracias - Thank you.